Human pre-miRNA Scramble Negative Control Expression Lentivector

Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| PMIRH000PA-1 | Human pre-miRNA Scramble Negative Control Expression Lentivector | Bacterial Streak | $792 |
|
||||
| PMIRH000VA-1 | Human pre-miRNA Scramble Negative Control Expression Lentivector | 1x10^6 IFUs | $954 |
|
||||
Overview
Overview
Supporting your studies with ready-to-go negative controls
No need to make a negative control for your Human Precursor miRNA Expression Lentivector Studies—SBI’s already built one for you. With the Human pre-miRNA Scramble Negative Control Expression Lentivector, you get the same lentivector that expresses our targeted miRNAs, but with a scrambled, non-targeting stem-loop sequence so you can easily control for increased miRNA effects.
The Human pre-miRNA Scramble Negative Control Expression Lentivector is available as both ready-to-transfect lentivector plasmid and ready-to-transduce pre-packaged lentivirus.
Why choose SBI’s precursor miRNA lentivectors?
Whether you’re using miRNAs to study cellular processes or develop the next generation of therapeutics, SBI’s comprehensive collection of Lenti-miRs™—human precursor miRNA lentivectors—are a superior alternative to synthetic RNAs. Like synthetic miRs, SBI’s Lenti-miRs can be transfected into target cells for transient miR expression. But unlike synthetic miRs, they can also be transduced into a variety of target cells—including primary cells, stem cells, and other hard-to-transfect cell lines—to create stable miR-producing cell lines, maximizing your options for miR expression.
In addition, SBI’s precursor miRNA lentivectors are designed for efficient expression and accurate processing into mature miRNAs, for native miR-like behavior. Each construct is cloned with the native stem-loop structure plus an additional 200 – 400 bps of upstream and downstream DNA, ensuring that the miRNA processing machinery operates just as it would with the native transcript.

And because our collection includes cancer and stem cell-related miRNA clusters, we offer a wider range of constructs than just the mature miRNA sequences listed in miRbase.
The precursor miRNA Expression Lentivectors drive expression of the miR precursor with the constitutive CMV promoter, for strong expression in common cell types including HeLa, HEK293, and HT1080 cell lines. Transductants/transfectants can be easily sorted using copGFP.
- Transfect or transduce the precursor miRNA lentivectors
- Create stably-expressing miR cell lines
- Leverage SBI’s highly-regarded lentivectors
- Obtain accurate miR processing and efficient expression through the use of native precursor sequences
- Use copGFP markers to identify desired clones
References
How It Works
How It Works
SBI’s Human pre-miRNA Expression Lentivectors use the native miRNA processing machinery

Supporting Data
Supporting Data
Get high levels of miRs from our Human pre-miRNA Expression Lentivectors
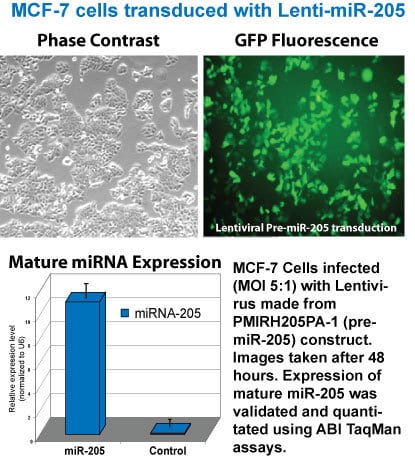
Lenti-miR-205 is expressed at high levels in MCF-7 cells
Lenti-miRs are overexpressed in HEK293 cells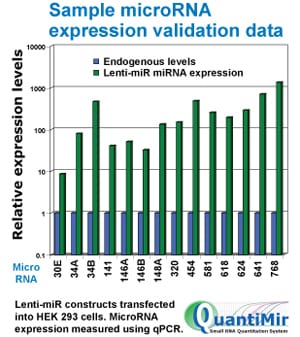
Lenti-miR stem cell cluster constructs are overexpressed 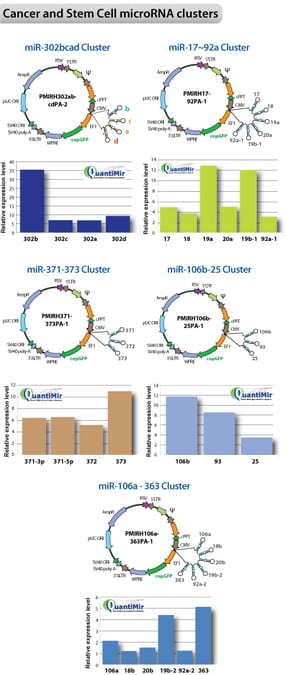
Lenti-miR stem cell cluster constructs are overexpressed 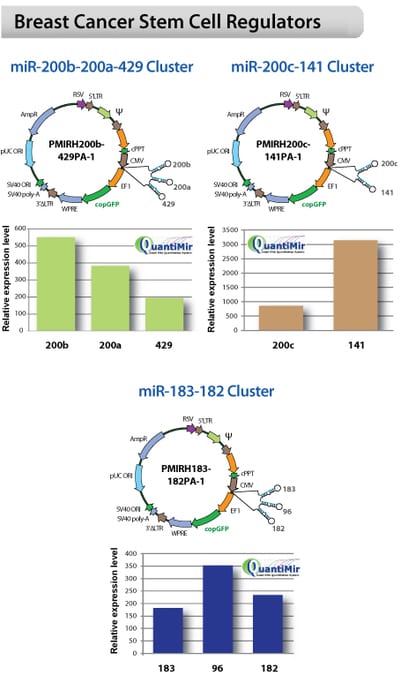
FAQs
Documentation
Citations
Related Products
Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| PMIRH000PA-1 | Human pre-miRNA Scramble Negative Control Expression Lentivector | Bacterial Streak | $792 |
|
||||
| PMIRH000VA-1 | Human pre-miRNA Scramble Negative Control Expression Lentivector | 1x10^6 IFUs | $954 |
|
||||
Overview
Overview
Supporting your studies with ready-to-go negative controls
No need to make a negative control for your Human Precursor miRNA Expression Lentivector Studies—SBI’s already built one for you. With the Human pre-miRNA Scramble Negative Control Expression Lentivector, you get the same lentivector that expresses our targeted miRNAs, but with a scrambled, non-targeting stem-loop sequence so you can easily control for increased miRNA effects.
The Human pre-miRNA Scramble Negative Control Expression Lentivector is available as both ready-to-transfect lentivector plasmid and ready-to-transduce pre-packaged lentivirus.
Why choose SBI’s precursor miRNA lentivectors?
Whether you’re using miRNAs to study cellular processes or develop the next generation of therapeutics, SBI’s comprehensive collection of Lenti-miRs™—human precursor miRNA lentivectors—are a superior alternative to synthetic RNAs. Like synthetic miRs, SBI’s Lenti-miRs can be transfected into target cells for transient miR expression. But unlike synthetic miRs, they can also be transduced into a variety of target cells—including primary cells, stem cells, and other hard-to-transfect cell lines—to create stable miR-producing cell lines, maximizing your options for miR expression.
In addition, SBI’s precursor miRNA lentivectors are designed for efficient expression and accurate processing into mature miRNAs, for native miR-like behavior. Each construct is cloned with the native stem-loop structure plus an additional 200 – 400 bps of upstream and downstream DNA, ensuring that the miRNA processing machinery operates just as it would with the native transcript.
And because our collection includes cancer and stem cell-related miRNA clusters, we offer a wider range of constructs than just the mature miRNA sequences listed in miRbase.
The precursor miRNA Expression Lentivectors drive expression of the miR precursor with the constitutive CMV promoter, for strong expression in common cell types including HeLa, HEK293, and HT1080 cell lines. Transductants/transfectants can be easily sorted using copGFP.
- Transfect or transduce the precursor miRNA lentivectors
- Create stably-expressing miR cell lines
- Leverage SBI’s highly-regarded lentivectors
- Obtain accurate miR processing and efficient expression through the use of native precursor sequences
- Use copGFP markers to identify desired clones
References
How It Works
How It Works
SBI’s Human pre-miRNA Expression Lentivectors use the native miRNA processing machinery
Supporting Data
Supporting Data
Get high levels of miRs from our Human pre-miRNA Expression Lentivectors
Lenti-miR-205 is expressed at high levels in MCF-7 cells
Lenti-miRs are overexpressed in HEK293 cells
Lenti-miR stem cell cluster constructs are overexpressed
Lenti-miR stem cell cluster constructs are overexpressed