pCDH-EF1α-CymR-T2A-Neo SparQ™ CymR Expression Vector
- Robust—increase expression up to 32-fold
- Adjustable—tune expression levels by titrating the amount of cumate
- Reversible—turn expression on, then off, then on again
- Versatile—choose from all-in-one formats that co-express CymR and your gene-of-interest, or two-vector systems where CymR is expressed from a different plasmid
- Powerful—suitable for in vivo applications

Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| QM400PA-2 | pCDH-EF1α-CymR-T2A-Neo (plasmid) | 10 µg | $693 |
|
||||
| QM400VA-2 | pCDH-EF1α-CymR-T2A-Neo (virus) | >1 x 10^6 IFUs | $743 |
|
||||
Overview
Overview
The repressor behind tight, titratable gene expression
With SBI’s SparQ™ Cumate Switch System, you can get inducible gene expression in mammalian cells through the binding of cumate, a non-toxic small molecule, to CymR. CymR can be delivered to cells on its own lentivector—such as with the pCDH-EF1α-CymR-T2A-Neo SparQ™ CymR Expression Vector (Cat.# QM400PA/VA-2)—or on the same lentivector as your gene-of-interest (Cat.#s QM800A-1 and QM812B-1).
- Robust—increase expression up to 32-fold
- Adjustable—tune expression levels by titrating the amount of cumate
- Reversible—turn expression on, then off, then on again
- Versatile—choose from all-in-one formats that co-express CymR and your gene-of-interest, or two-vector systems where CymR is expressed from a different plasmid
- Powerful—suitable for in vivo applications
With the SparQ System, expression levels are tightly controlled and increase with increasing cumate concentration until maximum induction is reached—see as much as a 32-fold increase in gene expression. Even better, induction is reversible, so you can turn expression levels on and off. Delivering negligible background expression in the absence of cumate, the SparQ System is an excellent choice for achieving controlled levels of gene expression.
The pCDH-EF1α-CymR-T2A-Neo Vector co-expresses CymR and a neomycin marker via the moderate EF1α promoter. Co-expression is mediated through a T2A element.

References
How It Works
How It Works
Tightly-controlled, inducible gene expression
SBI’s SparQ Cumate Switch System delivers robust, titratable gene expression with low background through three components:
- Cumate, a non-toxic, small-molecule inducer
- CymR, a repressor that binds to cumate operator sequences in the absence of cumate
- SparQ Lentivector that contains an MCS to clone-in your gene-of-interest, the cumate inducible promoter with cumate operator sequences (CuO) upstream of the MCS, and one or more markers
CymR has a high binding affinity for cumate and, as more cumate is added, fewer CymR molecules bind to the CuO sequences in the promoter resulting in increased expression. Exhibiting much lower background expression than similar systems, SBI’s cumate-inducible vectors can provide up to 32-fold induction of gene expression.

Supporting Data
Supporting Data
Tight expression control with low background with the SparQ Cumate Switch System
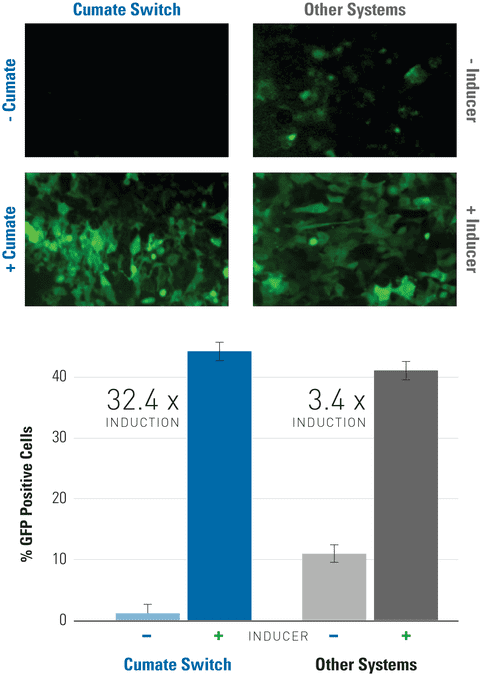
Figure 1. Get lower background and higher induction with the SparQ Cumate Switch System than other inducible systems.
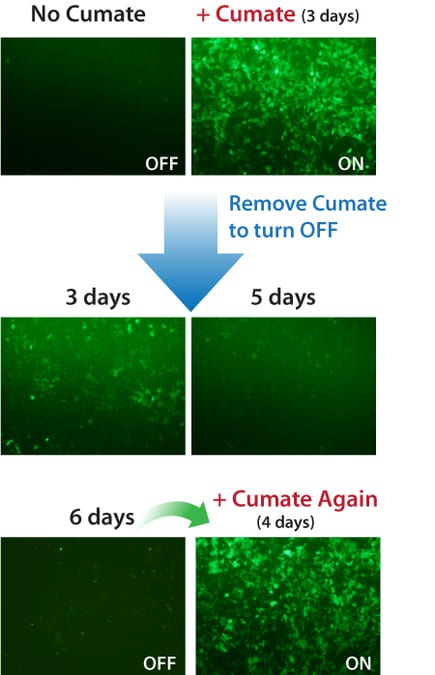
Figure 2. Gene expression with the SparQ Cumate SwitchSystem can be turned on and off, then on again.
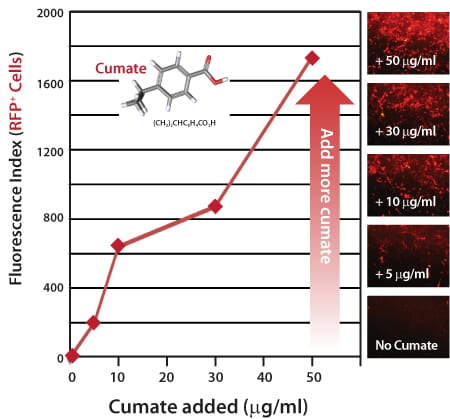
Figure 3. Gene expression with the SparQ Cumate Switch System is titratable, with increasing amounts of cumate inducing a linear increase in gene expression.
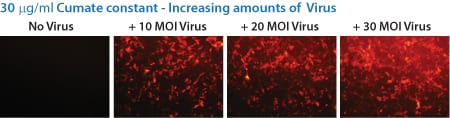
Figure 4. With the SparQ System, gene expression can also be titrated by increasing the amount of transduced SparQ lentivirus, even up to 30 MOI.
FAQs
Documentation
Citations
Related Products
Products
| Catalog Number | Description | Size | Price | Quantity | Add to Cart | |||
|---|---|---|---|---|---|---|---|---|
| QM400PA-2 | pCDH-EF1α-CymR-T2A-Neo (plasmid) | 10 µg | $693 |
|
||||
| QM400VA-2 | pCDH-EF1α-CymR-T2A-Neo (virus) | >1 x 10^6 IFUs | $743 |
|
||||
Overview
Overview
The repressor behind tight, titratable gene expression
With SBI’s SparQ™ Cumate Switch System, you can get inducible gene expression in mammalian cells through the binding of cumate, a non-toxic small molecule, to CymR. CymR can be delivered to cells on its own lentivector—such as with the pCDH-EF1α-CymR-T2A-Neo SparQ™ CymR Expression Vector (Cat.# QM400PA/VA-2)—or on the same lentivector as your gene-of-interest (Cat.#s QM800A-1 and QM812B-1).
- Robust—increase expression up to 32-fold
- Adjustable—tune expression levels by titrating the amount of cumate
- Reversible—turn expression on, then off, then on again
- Versatile—choose from all-in-one formats that co-express CymR and your gene-of-interest, or two-vector systems where CymR is expressed from a different plasmid
- Powerful—suitable for in vivo applications
With the SparQ System, expression levels are tightly controlled and increase with increasing cumate concentration until maximum induction is reached—see as much as a 32-fold increase in gene expression. Even better, induction is reversible, so you can turn expression levels on and off. Delivering negligible background expression in the absence of cumate, the SparQ System is an excellent choice for achieving controlled levels of gene expression.
The pCDH-EF1α-CymR-T2A-Neo Vector co-expresses CymR and a neomycin marker via the moderate EF1α promoter. Co-expression is mediated through a T2A element.
References
How It Works
How It Works
Tightly-controlled, inducible gene expression
SBI’s SparQ Cumate Switch System delivers robust, titratable gene expression with low background through three components:
- Cumate, a non-toxic, small-molecule inducer
- CymR, a repressor that binds to cumate operator sequences in the absence of cumate
- SparQ Lentivector that contains an MCS to clone-in your gene-of-interest, the cumate inducible promoter with cumate operator sequences (CuO) upstream of the MCS, and one or more markers
CymR has a high binding affinity for cumate and, as more cumate is added, fewer CymR molecules bind to the CuO sequences in the promoter resulting in increased expression. Exhibiting much lower background expression than similar systems, SBI’s cumate-inducible vectors can provide up to 32-fold induction of gene expression.
Supporting Data
Supporting Data
Tight expression control with low background with the SparQ Cumate Switch System
Figure 1. Get lower background and higher induction with the SparQ Cumate Switch System than other inducible systems.
Figure 2. Gene expression with the SparQ Cumate SwitchSystem can be turned on and off, then on again.
Figure 3. Gene expression with the SparQ Cumate Switch System is titratable, with increasing amounts of cumate inducing a linear increase in gene expression.
Figure 4. With the SparQ System, gene expression can also be titrated by increasing the amount of transduced SparQ lentivirus, even up to 30 MOI.